
生物信息 biological (Peter Godfrey-Smith and Kim Sterelny)
首次发表于 2007 年 10 月 4 日星期四;实质性修订于 2016 年 3 月 23 日星期三
自从 20 世纪 50 年代以来,信息的概念在生物学的许多领域中扮演着显著的角色。这种热情远远超出了那些似乎明显适用于该概念的领域,比如对感知、认知和语言的生物学研究,现在已经延伸到生物理论的最基本部分。调节生理系统的激素和其他细胞产物通常被描述为信号。关于基因在代谢过程和发育中起作用的描述通常以“转录”、“翻译”和“编辑”为基础。用于描述基因发挥作用的过程的最常用术语是“基因表达”。发育中的细胞命运是通过它们处理来自周围细胞和其他因素的“位置信息”来解释的。许多生物学家认为,有机体从卵到成体的发育过程可以理解为执行一个“发育计划”。其他生物学家则主张信息在进化中起到关键作用,而不是发育:例如,约翰·梅纳德·史密斯和埃尔斯·萨特玛里认为,进化中的重大转变取决于信息在世代间传递的数量和准确性的扩展。还有一些人认为,只有通过认识到信息“领域”与物质和能量领域并存,我们才能理解基因在进化中的作用。
哲学家和生物学家都为生物学中这种描述方式的地位进行了持续的基础性讨论。普遍认为,克劳德·香农所提出的并在数学信息理论中使用的信息概念是合法的、有用的,并且在生物学的许多领域中是相关的。从这个意义上讲,只要某物具有一系列可能的状态,并且一个变量与另一个变量的状态在物理上相关,那么它就是信息的源。但也有人认为,在生物学中使用信息语言的许多方式似乎使用了比香农的概念更丰富且更有问题的概念。一些人借鉴了心灵哲学中的目的语义传统来理解这个更丰富的概念。其他理论家则反驳说,香农的相关信息概念比它看起来更丰富。
少数派的传统认为,生物学中对信息的热衷是一个严重的理论错误,它培养了天真的遗传决定论,扭曲了我们对相互作用原因角色的理解,或者是一种隐含的二元论本体论。然而,这种怀疑的回应正在减弱,关键的怀疑者开始接受信息概念在生命科学中的适度但真实的作用。其他人认真对待了这一批评,但试图区分合理的信息诉求和误导性或错误的诉求。
1. 简介
生物学关注生物体——它们的结构、活动、空间和时间分布以及参与进化和发育历史。其中许多生物体从信息处理或表示的角度来看似乎更容易理解。这些包括感知、认知、信号传递和语言使用。我们不太关心这些认知背景下信息和表示概念的作用。在 20 世纪下半叶,生物学开始比以往更广泛地应用信息概念(及其相关概念)。对于许多生物学家来说,现在应该用信息的表达来理解生物体最基本的过程:对信号的响应、程序的执行和代码的解释。因此,尽管当代主流生物学是一个明显的唯物主义领域,它已经开始使用看似有意义或语义的概念。这些概念在唯物主义者(在某种程度上也包括其他人)中引起了长期的基础问题。
信息和其他语义概念的应用在遗传学以及与遗传学有密切联系的进化理论和发育生物学等领域中尤为显著。但信息语言不仅仅局限于这些领域:例如,激素通常被视为远距离信号,使一个器官系统能够与其他系统协调(Levy 2011)。话虽如此,引起最多讨论的用法是在描述基因与各种结构和过程之间的关系时出现的。对于许多生物学家来说,基因的因果作用应该从它们携带有关其各种产物的信息的角度来理解;也许还包括这些产物增强适应度的环境信息(Lorenz 1965; Shea 2013)。这些信息的表达可能取决于各种环境因素的存在,但对其他类型的信息也可以这样说。进一步分解,我们可以认识到基因有两个因果作用,因此在生物学中有两个潜在的解释作用。基因对于解释个体生物的发育以及解释特征在世代间的遗传都至关重要。信息在这两个解释背景下都被引用。
信息语言的一个重要用途是相对无争议的,通过基于关于 DNA 和 RNA 在细胞中制造蛋白质分子的作用的引人注目且确立的事实,这些事实总结在代表“遗传密码”的熟悉图表中,该图表将 DNA 碱基(C,A,T,G)的三联体映射到单个氨基酸,氨基酸是蛋白质分子的构建块。即使这种“编码”语言的使用也并非完全无争议(Sarkar 1996)。更重要的是,对信息语言在生物学中的使用的热情早于 DNA-RNA-氨基酸映射的发现(Schrödinger 1944),并且远远超出了这一映射,因此这种映射只是对生物学中信息概念采纳的部分解释。生物学中信息概念的当前应用包括:
将整个有机体表型特征(包括复杂行为特征)描述为基因中包含的信息所指定或编码的,
将细胞内的许多因果过程以及整个有机体发育序列可能以基因中存储的程序的执行方式来处理,
将基因(有时还包括其他遗传结构)的传递视为从父代到子代的信息流动。
为了进化理论化而将基因本身视为某种意义上的“信息制造”。信息成为生物世界的基本成分。
完全一般地描述理想化种群由于自然选择而发生变化的动力学。
关于这些观点的地位还没有达成共识。事实上,即使在描述动物间的交流时,使用信息概念也是有争议的,一些理论家否认这种交流是从一个动物到另一个动物的信息流动(Krebs 和 Dawkins 1984; Owren 等,2010)。这导致了生物学和生物哲学领域内日益增长的基础性讨论。一些人称赞信息概念的运用是一项重要的进展(Williams 1992)。其他人则认为几乎每一种生物学应用的信息概念都是一个严重的错误,它扭曲了我们的理解并导致了遗传决定论的持续存在(Francis 2003)。在这些极端观点之间,大多数可能的选择也得到了辩护。也许哲学界的大多数评论家都将他们的项目视为对当前各种信息描述进行分类的一种方式,区分合法的描述和非法的描述(Godfrey-Smith 2000; Griffiths 2001; Godfrey-Smith 2007; Shea 2013; Lean 2014)。哲学家们还试图对合法的描述给出还原论或自然主义的解释。
本条目按照以下方式进行。在下一节中,我们将讨论生物学中最常见的技术使用信息的问题,该问题依赖于香农和信息的数学理论。在这个背景下,我们介绍了一些更有争议的用法。在第 3 节中,我们讨论了最近试图从香农的技术概念中建立一个更丰富的概念的尝试。然后我们讨论了“遗传密码”在其原始意义上的地位(第 4 节),以及遗传学内外的信号系统(第 5 节)。然后我们看一下拒绝信息形式描述的方法(第 6 节)。关键的怀疑论观点是所谓的“平等论”观点:基因在进化和发展中起着极其重要的作用,但其他因素也起着作用,所以认为基因和仅仅基因指导或控制这些过程是错误的。最后两节讨论了个体生物发展通过执行一个程序进行(第 7 节),以及在进化的“重大转变”中理解遗传系统作用的信息使用(第 8 节)。
2. 香农的信息概念
在生物学中,通常以一个无争议但最简化的概念开始信息分析:因果或相关概念。空气中的烟雾与森林中的火灾相关。如果你在夜晚看到森林起火,你可以预测空气会有烟雾。如果你看到树上方升起的烟柱,你可以预测下方有火灾。相关性不必完美才能提供信息。总的来说,如果我们可以从信号中预测源的状态,那么信号就携带关于源的信息。这种信息意义上的概念与克劳德·香农(1948 年)相关,他展示了如何以一种有用的方式量化有关偶然性和相关性的事实,最初用于通信技术。对于香农来说,任何具有多种可能状态的事物都是信息的源,只要在特定场合上可能实现其中一种状态。任何其他变量如果其状态与源的状态相关,则包含关于该源的一些信息,或者携带关于该源的信息。这是一个程度问题;如果信号的状态更好地预测源,那么信号就携带更多关于源的信息;如果信号的状态预测能力较差,则携带的信息较少。
在这个意义上,信号可以携带关于源头的信息,而无需有任何生物系统来产生该信号,也没有任何生物系统在产生后使用它。当生物学家在描述基因作用或其他过程时采用这种信息的意义时,他们只是采用了一个量化的框架来描述这些系统内的普通相关性或因果关系。因此,信号携带关于源头的信息是一回事;而将生物过程解释为信号传递的结果则完全不同。这里有一个常见的例子是树木年轮。当树木形成年轮时,它建立了一个可以被我们用来推断过去的结构。年轮的数量和大小在香农的意义上携带着关于树木历史和环境的信息(这种技术,“树木年轮学”,在科学上非常重要)。尽管信息描述很有用,但在信息的术语中解释树木的行为方式是没有意义的。树木年轮中的信息只有人类观察者才能阅读或使用。树木本身并不利用年轮中的信息来控制其生长或开花。同样,我们可以注意到不同 DNA 序列在生物种群内部和种群之间的分布“携带着关于”这些种群之间的历史关系以及各个种群自身历史的信息。利用这些信息使得进化生物学家对生命历史有了更丰富和可靠的认识(关于这些用途的良好介绍可参见 Bromham 2008)。例如,有人认为与其他人类种群相比,非洲人群的线粒体 DNA 多样性更高,这是从非洲起源的人类相对较近期的扩散的指标。 在进行这些推断时,信息概念可以是有用的工具。但这只是树木年轮情况下的更复杂版本。对信息的引用具有推理用途,但绝不是解释性的。生物学中发现的大部分信息描述都具有这种特征。
然而,这两个类别之间的界限可能很难辨别。Steven Frank(2012)已经建立了自然选择引起的变化方程和信息论中使用的方程之间的详细映射。代际变化可以被看作是种群对其环境的信息积累。到目前为止,并没有声称种群以任何方式使用这些信息,情况类似于树木年轮:树木在其年轮中积累了关于其过去气候条件的信息,但树木对这些信息无动于衷。Frank 可能以更实质性的方式看待进化所获得的信息的作用:“选择使种群积累关于性状适应环境的信息”(2012:2391)。但是,对于这种信息,目前只能显示出推理作用,其他方面还存在疑问。
因此,哲学家们有时候通过说在生物学中有一种“信息”的类型,这种类型最初由香农描述,是无问题的,不需要太多哲学关注来引导讨论。有时候也会用术语“因果”信息来指代这个概念,这本质上是格赖斯在语言哲学中的“自然意义”(参见格赖斯的条目)(格赖斯 1957 年)中所使用的概念。在这个意义上,只要存在偶然性和相关性,就存在信息。因此,我们可以说基因包含有关它们所制造的蛋白质的信息,也可以说基因包含有关整个有机体表型的信息。但是当我们这样说的时候,我们所说的不过是当我们说烟和火之间存在信息联系,或者树环和树龄之间存在信息联系时所说的。更有争议的问题是生物学是否还需要另一个更丰富的信息概念。在这个更丰富的意义上,信息有时被称为“语义”或“意图”信息。
为什么我们认为生物学需要采用更丰富的概念呢?一个想法是基因在发育过程中扮演着特殊的指导作用,告诉胚胎如何生长。的确,基因携带着有关表型的香农信息:受精卵的基因组可以预测出很多最终的表型。例如,在哺乳动物中,染色体结构可以预测成年动物的性别。但是,如果基因与表型之间的信息关系被认为涉及一种独特的指令式因果模式,那么这不能是香农意义上的信息。对于环境因素来说,不仅仅是基因,都可以在香农意义上“携带信息”关于表型。在香农信息方面,环境和遗传原因的作用具有“平等性”(Griffiths 和 Gray 1994)。此外,如果我们使用香农的概念,那么基因型和表型之间的信息关系是对称的。例如,一旦读者知道本文的两位作者都是男性,他们就可以预测我们都携带着 Y 染色体。关于生物学中的信息的讨论与香农信息的这些特点是一致的,但也有一些不一致的地方。特别是,通常认为至少在一些将信息语言应用于基因的情况下,它们被赋予了一种不被赋予环境条件的属性,即使环境在预测上很重要。
此外,携带“语义信息”的消息往往被认为具有误传和准确传递的能力。存在误差的可能性。香农信息没有这个特点;如果我们使用香农的术语,我们不能说一个变量“携带了关于另一个变量的错误信息”。但是生物学家显然希望在谈论基因时使用这种语言。基因携带着一条被认为应该被表达的信息,无论它是否实际被表达。因此,20 世纪 60 年代沙利度胺引起的发育障碍是由于无法使用应该控制肢体发育的遗传信号。这些基因存在并且活跃;沙利度胺并没有通过引起突变来造成伤害。
这些通常是文献中用来显示引入了比香农更丰富的信息概念到生物学中的“标志”。但是,在信息概念的应用中,较少争议和较多争议的关键区别在于,在较丰富的情况下,信息的使用被认为有助于解释生物系统如何完成其功能-细胞如何工作,一个卵子如何发育成为成体,遗传继承机制如何使复杂表型的进化成为可能。
在这一点上,桌上有许多选择。其中之一是否认基因、细胞和其他生物结构在解释其行为的方式上确实传递信息,但仍然主张这是一个有用的类比或模型。这个想法是,信息和表示系统的典范案例(认知复杂的代理人,彼此思考和交流)与生物系统之间存在有用的相似之处。例如,激素可以有用地被视为信息,因为它们是小巧、稳定、能量消耗低的物质,可以在不衰变的情况下长距离传播(相对于它们自身的大小),到达特定的位置后产生可预测、具体和重要的效果。但是,虽然这是一种有帮助的思考方式,也许我们不应该过于认真地谈论信息传递。例如,我们不应该把关于催乳素是对怀孕的报告还是对乳腺的指令的问题当作有确切答案的问题来对待。Arnon Levy 提出了生物信息的最复杂版本(Levy 2011)。
第二个选择是主张基因和其他生物结构实际上携带语义信息,而它们的信息特性解释了这些结构在生物过程中的独特作用。如果我们将基因或细胞视为实际携带语义信息,我们面临的问题就改变了;谁或什么可以被视为构成或阅读这些信息?具有语义信息的典型结构案例——图片、句子、程序——是由智能体的思想和行动构建的。因此,我们需要展示基因和细胞如何能够携带语义信息(它们既不是智能系统本身,也不是智能的产物),以及它们所携带的信息如何解释它们在生物学中的作用。我们需要某种关于语义信息的还原解释(可以说,我们也需要这样的解释来理解认知)。我们可以在自然主义的心灵哲学中寻找这样的分析方法。
第三个选择是主张因果信息本身可以解释生物现象,不需要额外的信息概念。在这种观点下,生物系统可以适应发送或接收携带因果信息的信号。举例来说,催乳素的升高与妊娠是相关的,并非偶然。同样,乳汁的产生是对乳腺中这些升高水平的注册的设计反应。虽然乳汁分泌对催乳素水平具有同样多的因果信息,就像这些水平对乳汁流动具有的信息一样,但在源头和接收器之间存在着物理上的不对称性,因此存在方向性。(出生后,哺乳刺激垂体腺产生催乳素,因此物理上的不对称性是从垂体腺向乳房传递妊娠信号,使其适应哺乳)。布莱恩·斯凯姆斯(Brian Skyrms)的工作在将因果信息重新置于中心舞台上非常重要,部分原因是他的工作表明,发送者-接收者信号系统不必是认知复杂的代理人(Skyrms 2010)。发送和接收因果信息可以在简单系统中出现和稳定,当然也可以在不比细胞更复杂的系统中出现。
3. 目的语义学和其他更丰富的概念
如上所述,一些哲学家和生物学家认为,关于基因的大部分信息论讨论使用了比香农的更丰富的概念,但这个概念可以进行自然主义分析。目的是理解基因在语义上指定其正常产物的想法,类似于一些典型的符号现象中所见到的那种意义。
如果将基因视为在这个意义上“传递信息”,那么这个信息显然具有规定性或命令性的内容,而不是描述性或指示性的内容。它们与其效果的“适应性”是这样的,如果基因和最终产生的结构(表型)不匹配,我们所面对的是未实现的指令,而不是不准确的描述。或者,也许可以认为基因在告诉发育中的表型它可以期待的环境。那些基因来自的基因库已经经过选择筛选。在澳大利亚的干旱地区,那些有助于发展形状和表面限制水分流失的叶子的基因已经变得普遍。因此,也许我们可以将这些基因视为告诉树木:条件将会干燥(有关这一思路,请参见 Shea 2011, 2013)。
在理解这些观点时,通常的做法是利用一个丰富的生物学功能概念,其中实体的功能源于自然选择的历史(参见生物学中的目的论概念)。这种做法在心灵哲学中是常见的,因为在解释心理状态的语义属性时也会出现类似的问题(Millikan 1984)。当一个实体经历并受到自然选择的塑造时,这可以为该实体的因果能力提供一种目的性或规范性描述的基础。使用标准例子(Wright 1973),心脏的功能是泵血,而不是发出重击声音,因为正是前者的效果导致了心脏受到自然选择的青睐。希望通过类似的“目的功能”策略来理解基因的语义属性,或许还有其他具有语义属性的生物结构。
关于遗传信息的目的语义学方法的概念最早由 Sterelny 等人(1996)提出;另请参阅 Maclaurin(1998)。杰出的生物学家约翰·梅纳德·史密斯(John Maynard Smith)采取了类似的方法,试图理解他自己对生物学中信息概念的热情(Maynard Smith 2000;另请参阅随后的评论)。Eva Jablonka 也捍卫了这个想法的一个版本(Jablonka 2002)。她的处理方式更加非传统,因为她试图将环境信号视为具有语义信息的一部分,与基因一起,如果它们被生物体以适当的方式使用。Nick Shea 迄今为止开发了最复杂的基因目的语义学处理方法(请参阅 Shea 2007a,b,2011,2013)。
发展这些想法的一种方式是关注遗传机制作为一个整体的进化功能(Godfrey-Smith 1999)。Carl Bergstrom 和 Martin Rosvall 采取了这种方法,强调了代际基因流的适应性和令人印象深刻的工程特征,以发展他们的“信息传递意义”。Bergstron 和 Rosvall 指出,代际基因流的结构使得任意序列的传输成为可能(因此,信息在媒介中相对不受限制);信息被紧凑而稳定地存储;带宽大且可无限扩展。DNA 序列可靠地以非常高的保真度进行复制,因此传输是准确的,然而 DNA 和氨基酸序列之间的映射似乎经过优化,以减少发生的错误的影响。许多点突变被映射到相同或化学相似的氨基酸上。Bergstrom 和 Rosvall 得出的结论是,DNA 复制是一个精心设计的信息通道,并且我们可以从 DNA-氨基酸系统本身的特性中得知这一点。为了知道 DNA 复制的特性是由其信息传递能力解释的,我们不需要知道父代向子代传递的信号是什么(Bergstrom 和 Rosvall 2011)。
更常见的路线,也是 Sterelny 等人(1996 年)和 Maynard Smith(2000 年)所采取的路线,是专注于特定遗传元素的自然选择。这种观点面临着一个直接的问题,因为特定的遗传元素(或整个遗传系统)具有进化功能显然不足以使基因携带语义信息。腿是用来走路的,但它们并不代表走路。酶是用来催化反应的,但它们并不指示这种活动。腿和酶应该做某些事情,但这并不意味着它们成为信息载体,在一个丰富的超过 Shannon 的意义上。为什么基因会这样呢?Sterelny、Smith 和 Dickison(1996 年)提出,基因与腿的区别在于基因已经被选择来在发育过程中发挥因果作用。然而,他们补充说,任何具有类似发育作用并被选择来发挥该作用的非遗传因素也具有语义属性。因此,Sterelny、Smith 和 Dickison 希望将非常丰富的语义属性归因于基因,而不仅仅是基因。一些非遗传因素具有相同的地位。即便如此,根据这种观点,许多相当合理的情况都不是信息化的:催乳素,像大多数激素一样,没有特定的发育功能,因此它不会被视为向乳腺传递信息。这是为什么 Levy 将信息的讨论视为纯粹的隐喻的原因之一(Levy 2011)。与 Sterelny、Smith 和 Dickison 的观点相反,约翰·梅纳德·史密斯(John Maynard Smith)在他 2000 年的论文中认为,只有基因才能携带有关表型的语义信息。他提出,与其他发育资源相比,适应性基因与表型结果之间的关系是任意的;基因-特征关系就像词-物体关系一样。这个想法很有趣,但很难具体化。 一个问题是,如果通过许多中介环节进行操作,任何因果关系都可能看起来“随意”,因为在这些环节上有许多可能的干预措施,这些干预措施会改变因果链的产物。关于随意性的问题在第 4 节中进一步讨论。
尼古拉斯·谢亚为了区分普通的生物功能和表征功能,借鉴了鲁思·米利坎的目的语义理论中更为复杂的机制。对于米利坎来说,任何具有语义属性的对象都扮演着一种在两个“协作设备”之间进行中介的角色,即生产者设备和消费者设备。在指示信号或表征的情况下,表征被认为会以某种方式影响消费者的活动,只有在某种情况下,这种影响才会进一步促进消费者的生物功能的表现。在命令性表征的情况下,表征被认为会通过诱导消费者来实现某种情况。谢亚将遗传信息视为具有指示性和命令性内容,并依赖于生产者和消费者(谢亚 2007b,2011,2013)。谢亚在强调生产者和消费者的角色方面遵循米利坎的观点,而贾布隆卡(2002)则试图仅通过强调消费者机制来实现尽可能多的功能。显然,关注生产者和消费者机制是这个讨论的一步前进,但我们将在第 5 节中看到,它也带来了问题。在思考继承和发展时,目前尚不清楚是否存在可以独立识别的机制,这些机制可以被视为发送者和接收者、生产者和消费者。
通过遗传语义方法构想的整体图景具有不可否认的吸引人的结构特征。如果这个计划成功,我们将会有一个无争议的关于信息的感知,通过香农的方式,适用于各种物理相关性。然后,通过识别一些这些信号已被利用或产生来驱动生物过程的情况的子集,可以进一步发展这个图景。此外,也许我们可以在具有正确类型的自然选择历史的情况下,诉诸于丰富的语义属性,以解释基因在发育中的独特作用,也许还有其他因素。基因和少数非遗传因素将具备这些属性;大多数在生物发育中具有因果作用的环境特征则不具备。虽然仍然存在许多细节问题,但总体图景的吸引力至少为一些人提供了沿着这些思路坚持下去的充分理由。
4. 遗传密码
到目前为止,我们主要讨论了信息的概念;并没有太多关于“编码”的讨论。除了我们对 Bergstrom 和 Rosvall 关于信息的“传输意义”的讨论之外,到目前为止的讨论并没有强调基因所涉及的细胞层面过程的独特特征,比如“遗传密码”的类似语言的组合结构。即使在关注发育和遗传的情况下,DNA 的这些结构特征及其与氨基酸的关系并不是生物学中关于信息的一些观点的核心。如上所述,对生物结构进行语义化描述的热情可以追溯到遗传密码被发现之前(详见 Kay 2000 的详细历史处理)。但是文献中的一种思路与上述思想重叠,专注于遗传机制的特殊特征,以及将“遗传编码”作为这些机制的一种偶然特征的观念。
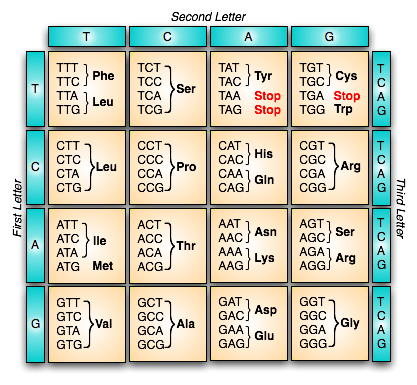
Peter Godfrey-Smith 和 Paul Griffiths 都认为,在遗传学中有一种非常受限制的使用方式,这是合理的(Godfrey-Smith 2000; Griffiths 2001)。这个想法是基因通过细胞内的“转录和翻译”机制,以一种特殊的方式“编码”蛋白质分子的氨基酸序列。基因通过一种模板化过程来指定氨基酸序列,该过程涉及两种完全不同类型的分子(核酸碱基和氨基酸)之间的规则映射。这种映射规则是组合的,而且显然是任意的(在一种难以精确描述的意义上,尽管参见 Stegmann 2004,讨论了这个想法的不同版本)。
下面的图 1 总结了标准遗传密码。诸如“Phe”和“Leu”之类的三字母缩写是氨基酸分子的类型。
图 1. 标准遗传密码
这种对基因信息特性的非常狭窄理解基本上与弗朗西斯·克里克(1958 年)的早期提议相一致。论点是,这些低级机械特性使基因表达成为一个具有重要类比于典型符号现象的因果过程。
有人认为,一旦我们从简单的原核生物(细菌和古菌)的遗传学转向真核细胞中的遗传学(Sarkar 1996),这种类比就变得值得怀疑。主流生物学倾向于将在真核生物中出现的复杂性视为仅仅是不影响我们对基因表达工作方式的基本图景的细节(对于那些认为这些复杂性真的很重要的人来说,可以参考(Griffiths 和 Stotz 2013)的广泛讨论)。一个例子是 mRNA 转录本的编辑和“剪接”。基因表达的初始阶段是使用 DNA 进行模板过程,构建一个中间分子 mRNA 或“信使 RNA”,然后将其用作蛋白质制造的模板。蛋白质是通过将一系列氨基酸分子串联在一起制成的。在真核生物中,mRNA 在使用之前通常会被广泛修改(“编辑”)。此外,真核生物中的大部分 DNA 根本不进行转录和翻译。其中一些“非编码”DNA(再次注意信息化语言)肯定是功能性的,用作与 DNA 结合的蛋白质的结合位点,从而调节转录的时机和速率。完全无功能的 DNA 的范围仍不清楚。这些事实使得真核生物的 DNA 对蛋白质的氨基酸序列的预测变得不那么直接,但可以认为这并不太影响激发引入符号或语义描述模式的基因表达机制的关键特征。
因此,Godfrey-Smith(2000)和 Griffiths(2001)的论点是,基因和只有基因具有一种信息或语义属性:编码蛋白质分子的氨基酸序列。但这种关系“延伸”的范围仅限于氨基酸序列。它并不能证明基因编码整个有机体表型的观点,更不用说为生物学中信息或语义语言的广泛使用提供基础。基因当然可以在产生整个有机体表型的过程中发挥可靠的因果作用。但如果要用信息术语来描述这种因果关系,那么它就是普通的香农信息,同样适用于环境因素。也就是说,可以争论基因作用的特异性以及染色体上给定位点的一系列实际和可能的替代品意味着基因对表型具有细粒度的因果控制,并且很少有其他发育资源具有这种形式的因果控制(Waters 2007; Maclaurin 1998; Stegmann 2014)。相比之下,Griffiths 和 Stotz(2013)认为这些其他因素通常是“Crick 信息”的来源,因为它们有助于指定基因产物的线性序列。我们将在第 6 节中回到这个问题。
基因编码氨基酸序列的想法最吸引人,但也可能存在问题,其中之一就是所谓的基因密码的“任意性”。任意性的概念在其他关于遗传信息的讨论中也起到了作用(Maynard Smith 2000; Sarkar 2003; Stegmann 2004)。人们常说标准的基因密码具有任意性特征,因为如果在“翻译”基因信息的机制中有补偿性的变化,那么 DNA 碱基三联体与氨基酸之间的许多其他映射也是可能的。弗朗西斯·克里克提出,基因密码的结构应该被视为一个“冻结的意外”,最初是高度偶然的,但现在对进化来说很难改变(Crick 1958)。但是,任意性的概念和冻结意外的假设已经变得有问题。首先,正如我们在上面讨论 Bergstrom 和 Rosvall(2011)时指出的那样,该密码并不是以许多其他密码同样有效的意义上的任意性。相反,现有的密码在最小化错误成本方面接近最优。从概念上讲,如果任何因果关系通过许多中介环节进行操作,它似乎都可以看作是“任意的”。每个分子结合事件发生的机制并不“任意”。使密码看起来任意的是碱基三联体与氨基酸之间的映射是通过一个具有多个中介环节的因果链介导的(尤其涉及“转运 RNA”分子和将氨基酸与这些中介分子结合的酶)。因为我们经常关注 DNA 和蛋白质之间的“长距离”连接,所以因果关系看起来是任意的。如果我们关注任何其他生物级联中相隔三到四个中介环节的步骤,因果关系看起来也同样“任意”。因此,任意性的概念本身就是难以捉摸的。 而且根据实证,标准遗传密码的结构比人们曾经想象的更加系统和非偶然(Knight,Freeland 和 Landweber 1999)。任意性的概念也被用于讨论基因和表型之间联系的更一般意义上。Kirscher 和 Gerhart(2005)讨论了一种源于蛋白质分子细节及其与基因调控的关系的任意性。调控基因作用的蛋白质往往具有独特的结合位点,进化可以独立改变这些位点。为了结合,蛋白质必须能够附着到位点上,但这仅需要与蛋白质的局部特征相一致,而不是严格限制其整体形状(Planer 2014)。这导致了一系列可能的基因调控过程。因此,在讨论某些生物因果性的信息性质时,人们经常倾向于诉诸任意性的观念。
5. 信号系统
在第 2 节中,我们注意到 Brian Skyrms 关于信号系统的研究使得这个框架非常适合于相当大范围的生物现象。正如我们上面所提到的,将胰岛素、睾酮和生长激素等激素视为信号是非常直观的,因为它们在身体的一部分产生,并传播到其他部位,在那里它们与“受体”相互作用,以修改各种其他结构的活动。将激素描述为“化学信息”是常规做法。Skyrms 的框架之所以适用于这些设计的因果关系和响应系统,有三个原因。首先,正如前面所提到的,这个框架表明,信号传递不需要智能或对信号含义的智能把握。
其次,信号传导模型中最简单的情况是存在共同利益的情况。发送者和接收者受到相同结果的利益或不利影响。虽然完全的共同利益在生物体间的通信中并不常见,但生物体内的细胞和其他结构共享着共同的命运(有复杂的例外情况)。因此,在这方面,基本模型可能更适用于器官间的通信,而不是生物体间的现象。
第三,在许多这些生物系统中,作为信号系统的抽象结构——环境源、发送者、消息、接收者、响应——在具体的生物机制中非常自然地映射。例如,Ron Planer 最近提出,我们应该将基因表达视为信号系统的运作(Planer 2014)。他的观点非常细致,因为发送者、接收者和消息的身份在不同情况下有所变化。例如,当蛋白质是转录因子时,基因被视为发送者。他对待其他基因-蛋白质关系的方式不同。他的观点的细节在这里并不重要。重点是细胞中存在着一些机制——基因、蛋白质 mRNA 转录本、核糖体及其相关的 tRNA——可以合理地映射到发送者-接收者系统上。将信息处理的发送者-接收者结构强行映射到细胞的分子机制上并不是一种迫使。
然而,虽然这个框架非常自然地适用于细胞内和细胞间的过程,但其他建议如何与这个框架自然地结合起来则不太清楚。例如,在 Bergstrom-Rosvall 关于代际传递的观点中,发送者和接收者是谁或是什么?也许在多细胞生物的情况下,接收者独立于并先于信息存在。因为在受精核开始基因表达之前,卵子就是一个复杂且高度结构化的系统,而这个结构在引导基因表达中起着重要作用(Sterelny 2009; Gilbert 2013)。但大多数生物是单细胞原核生物,当它们分裂时,并不明显存在一个在代际遗传信息之前独立存在的子代。
同样,尼古拉斯·谢的米利坎派生分析似乎不能自然地映射到可独立指定的生物机制上。对他来说,遗传信息的发送者是自然选择对人群的基因库进行操作和过滤;信息由整个有机体的发育系统阅读(Shea 2013)。但是,发送者-接收者或生产者-消费者框架与独立认可的生物机制越不明确,对该案例进行虚构主义或类比分析就越有可能。因此,我们可以看到读者作为整个发育系统和读者作为细胞膜上的受体之间存在重要差异。转录和翻译的细胞水平机制(尤其是核糖体/tRNA 机制)确实是核酸序列的读者或消费者,其功能是在细胞的其他位置产生各种用途的蛋白质产物。但是,这种因果关系模式的实现仅适用于细胞水平,在该水平上,转录和翻译装置显示为机器的一部分。发育过程中最非凡的特征之一是它在没有整个有机体发育的中央控制的情况下可靠且可预测地进行。例如,没有任何机制会检查左侧肢体是否与右侧肢体大小相同,以确保对称性。同样,存在 DNA 序列读取器和一些具有明确“读者”的细胞间发送者-接收者系统(如神经结构),但没有解释胚胎中整个肢芽结构的消息的更高级别读者。
6. 生物学中对信息概念的拒绝
一些生物学家和哲学家认为,信息和语义概念的引入对生物学产生了不良影响,它培养了各种解释性幻觉和扭曲,也许还伴随本体论混淆。在这里,我们将调查一些更强调这种观点的主张,但在许多其他讨论中也可以察觉到一定程度的不安(例如,参见 Griesemer 2005)。
被称为发育系统理论(DST)的运动经常反对生物学中信息概念的主流用法,主要是因为这些概念扭曲了我们对基因所涉及的因果过程的理解。有关这一话题的开创性讨论,请参见 Oyama(1985),还有 Lehrman(1970);Griffiths 和 Gray(1994);Griffiths 和 Neumann-Held(1999)。这些理论家对生物学中信息概念的使用有两个相关的反对意见。一个是信息模型是预形成主义的观念。预形成主义在其原始形式中实际上将发育简化为生长:在受精卵内存在着一个未来成体的微型形式。预形成主义不能解释一个有组织、有区别的成体是如何从一个组织较少、更均匀的卵中发育出来的;它否认了这一现象。DST 的支持者怀疑发育的信息模型也是如此。例如,假设“语言器官”的指令编码在新生婴儿的基因组中,你并没有解释语言能力如何在一个缺乏语言能力的有机体中发展起来,你培养了这样一个幻觉,即不需要这样的解释。(参见 Francis 2003,对于信息的诉求导致生物学中的伪解释这一观点的特别有力版本。)
其次,生物信息理论家经常支持“平等论”:基因在发育中起着不可或缺的作用,但其他因果因素也同样重要,并且没有理由将基因对发育的贡献置于特权地位。这一观点通常以引用理查德·勒温汀(Richard Lewontin)关于发育相互作用的复杂性和上下文敏感性的论证来支持,以及他因此而提出的我们通常无法将基因和环境对特定表型结果的因果责任进行正常分割的论证(Lewontin 1974, 2000)。生物信息理论家认为,基因和基因作用的信息模型很容易忽视平等性,并将某种因果优先性归因于这些因素,尽管它们只是问题过程中一组必不可少的贡献者之一。一旦将复杂系统中的一个因素视为信息性质,其他因素往往被视为纯粹的背景,作为支持而不是真正的因果行动者。人们自然而然地认为基因指导、控制或组织发育,其他因素提供必要的资源。但是,论证认为,在生物系统中,基因的因果作用与许多其他因素的作用紧密相互关联(通常松散地被归为“环境”)。有时,一个基因对广泛的环境背景都会产生可靠的影响;有时,一个环境因素对广泛的基因背景都会产生可靠的影响。有时,无论是基因还是环境原因在其操作中都高度依赖上下文。保罗·格里菲斯(Paul Griffiths)强调了这个问题,他认为描述基因的信息模式可以促使上下文独立性的出现:
基因是指令-它们提供信息-而其他因果因素只是物质.... 一个同性恋基因是一个指令,即使(因为其他因素)这个人是异性恋。(格里菲斯 2001:395-96)
声称与信息或语义概念的使用相伴的推理习惯和关联使我们认为基因具有额外和微妙的额外因果特异性。即使人们愿意公开接受复杂生物系统中(大多数)原因的依赖于上下文,这些习惯也会产生影响。因此,DST 理论家认为,将基因和仅基因视为携带表达在其效应中的“信息”的做法是误导性的。这样说几乎不可避免地将环境因素视为次要参与者。
平等论已经成为了广泛讨论和回应的焦点。在一篇有帮助的论文中,乌尔里希·斯特格曼(Ulrich Stegmann)表明,平等论实际上是一系列论点而不是单一论点(Stegmann 2012)。对平等论的解释方式有些使这个想法变得毫无争议,仅仅是对发育的复杂和互动性的坚持,或者指出基因与非遗传因素一样,有稍微不同的版本,具有稍微不同的效应(保持其他因素恒定)。由于营养环境、垃圾位置和出生顺序,基因上的表观遗传标记也可能有稍微不同的变体,具有稍微不同的效应。其他版本的主张则更具争议性。
对于平等论的一个回应是接受这样一种观点,即基因只是一组个别必要且集体充分的发育因素之一,但仍然主张基因在发育中发挥着独特且特别重要的作用(Austin 2015; Lean 2014; Planer 2014)。如上所述,沿着这个方向最有希望的建议可能是基因对发育施加了一种普遍、普遍和细粒度的因果控制。每个生物体的表型的许多特征以稍有不同的版本存在,这是由于相关基因的等位变异的结果。没有其他发育因素能够施加类似于基因的普遍、普遍和细粒度的控制(Woodward 2010; Stegmann 2014)。因此,Stegmann 通过对比干预细胞活性的聚合酶数量和干预 DNA 模板本身的效果,阐述了 Woodward 的观点,即基因对表型具有特定的控制作用。聚合酶在因果上至关重要,但改变其浓度只会修改合成速率,而不会改变产生的序列。而对 DNA 序列本身的修改并非如此,因此 DNA 序列比聚合酶更具因果特异性。
Shea 采取了不同的方法,认为不同的因果因素具有不同的进化历史。一些因果因素只是环境的持续特征(例如重力)。其他因果因素是发育中的生物体由于选择历史而经历的。例如,洞穴确保蛋和雏鸟在相对恒定的温度和湿度下发育。但洞穴并不是自然选择的遗传机制。它们并不是为了确保海鸟雏鸟与父母相似而存在的。相比之下,一些其他发育特征之所以存在并在发育中起作用,是因为它们在选择历史中具有选择优势,这些机制有助于确保父子相似性。Shea 认为,基因,可能是基因上的表观遗传标记,以及可能还有其他一些发育资源,都受到这种形式的自然选择的塑造。因此,基因和可能还有其他一些发育因素在发育中起着独特的作用,尽管许多其他因素在因果上是必要的(Shea 2011)。
总之,对于在思考发展过程中使用信息术语需要谨慎有充分的理由。但是,我们也可能过高估计了信息化发展观念与基因在发展中发挥独特重要作用之间的联系的强度。有一些方法可以辩护基因发挥特殊作用的观念,同时承认发展的互动性质。此外,对信息概念的雄心勃勃的运用并不仅限于主流生物学思维中的人士。Eva Jablonka 和 Marion Lamb 在维护相当异端的遗传和进化观点时,基于信息概念,包括对“拉马克”思想的倡导(Jablonka 和 Lamb 2005)。他们认为,信息描述的一个有用特征是它们允许我们以共同的货币比较不同遗传系统的属性。此外,本文的一位作者使用信息概念区分了基因与其他遗传因素的进化作用,同时证明了非遗传性的进化重要性(Sterelny 2004, 2011)。因此,以各种方式,信息观点可能有助于讨论非正统的理论选择,包括非基因遗传机制。
7. 生物信息程序
谈论基因“程序”在生物学的普及介绍和生物学本身中都很常见。通常,这个想法只是一种生动(但可能是误导性的)方式,用来引起对发育的有序、良好控制和高度结构化特征的关注。总体而言,尽管细胞内和细胞间相互作用的复杂性非常高,而且发育发生的物理环境永远无法精确控制,但发育的结果令人惊讶地稳定和可预测。因此,当生物学家们谈论“程序性细胞死亡”时,他们同样可以说,在一类重要情况下,细胞死亡是可预测、有组织和适应性的。
有人试图在计算系统和生物发展之间建立更紧密和有启发性的类比。特别是,罗杰·桑森(Roger Sansom)一直在努力发展生物发展和连接主义计算模型之间的紧密和有启发性的类比(Sansom 2008b,a,2011)。这种观点的优点在于承认发展没有中央控制;生物体的发展是由细胞内部和细胞之间的局部相互作用的结果。然而,对于程序开发类比的最有希望的想法似乎是指细胞内部过程与现代计算机的低级操作之间的明显类比。细胞内部的一个关键因果过程是基因网络中的级联上调和下调。一个基因会产生一个与另一个基因结合并因此下调的产物,然后阻止该基因产生上调另一个基因的产物...依此类推。我们在这里有一系列事件,通常可以用变量之间的布尔关系来描述。一个事件可能只是另外两个事件的合取,或者是它们的析取。下调是一种否定,网络中可能存在双重和三重否定。基因调控网络通常具有足够丰富的这种结构,以便将其视为一种计算。计算机芯片的“与门”,神经元的“与门”和基因的“与门”具有真正的相似之处。
在谈论信号网络而不是程序时,Brett Calcott 表明,发育中的果蝇胚胎的位置信息取决于这种布尔结构,肢芽发育取决于分化为肢芽的细胞整合一个正信号和两个负信号,因此肢芽在胚胎的前中线上以规律的模式发育。Calcott 表明,用这些信号网络及其布尔结构来思考发育具有真正的解释价值,因为它使我们能够解释位置信息如何在进化中容易被重复使用。果蝇翅膀上的斑点可以很容易地进化,因为告诉细胞它们在翅膀上的位置的网络已经存在,所以翅膀斑点的进化只需要一个简单的突变改变将位置信息与色素产生联系起来(Calcott 2014)。Ron Planer 认为基因调控具有这种布尔结构,并且我们可以将每个基因表示为实例化的条件指令。条件的“if”部分指定了打开基因的分子条件;条件的“then”部分指定了由基因制造的氨基酸序列。与 Calcott 一样,Planer 指出,这些条件指令可以并且经常被链接在一起构建复杂的控制网络。即使如此,Planer 认为,虽然这些是信号网络,但不应将其视为计算机程序。例如,指令的组合没有固有的顺序;我们可以将每个基因表示为特定的条件指令,但指令集本身没有告诉我们从何处开始和结束(Planer 2014)。
8. 生物信息和进化
生物信息也成为了进化过程的普遍讨论焦点,特别是与遗传机制相关的信息。这个讨论中的一个观点误解了信息及其在生物过程中的作用。特别是 G.C.威廉姆斯认为,通过对基因在进化中的作用的反思,我们可以推断出存在一个信息的“领域”,它存在于物质和能量的物理领域之外(威廉姆斯,1992 年)。理查德·道金斯支持类似的观点,认为进化的长期路径由继承信息的渐变组成,就像一条“在时间中流动而非空间中流动”的河流(道金斯,1995 年:4)。这是对更常见观点的延伸,即存在着“信息基因”,应该将其理解为与由 DNA 构成并在空间和时间上定位的“物质基因”不同的东西(海格,1997 年)。认为存在两种不同的东西是错误的;即存在一个物理实体——一串碱基,和一个信息实体——一条信息。确实,对于进化(以及许多其他)目的来说,基因通常最好从其碱基序列(C、A、T 和 G 的序列)的角度来考虑,而不是从其完整的物质属性集合的角度来考虑。这种思维方式本质上是一种抽象(Griesemer,2005 年)。我们正确地忽略了 DNA 的某些属性,而关注其他属性。但是将这种抽象视为一个额外的实体,并与物理领域有神秘的关系是错误的。
将信息性思想与进化理论中的一般问题联系起来的其他方法似乎更有前途。正如约翰·梅纳德·史密斯(John Maynard Smith)、埃尔斯·萨斯玛里(Eors Szathmáry)、马克·里德利(Mark Ridley)和理查德·道金斯(Richard Dawkins)以不同的方式强调的那样,导致重要进化结果的遗传机制必须满足一些相当特殊的条件(Dawkins 1995;Jablonka 和 Szathmáry 1995;Szathmáry 和 Maynard Smith 1997;Ridley 2000)。例如,梅纳德·史密斯和萨斯玛里声称,遗传系统必须在产生新组合的能力上是无限的或“不确定的”,但同时也必须保持高度的传递保真度。关于遗传系统与生物结构之间的这一事实通常被认为揭示了解释生命起源中最紧迫的问题之一。如果繁殖依赖于关键成分的复制来启动新一代(无论我们是否将这些成分视为指令),那么这些成分必须被准确地复制。然而,准确的复制显然依赖于复杂的分子和细胞内机制,而这些机制本身是长期适应性进化的结果,因此依赖于深层次的生命系统。那么繁殖是如何开始的呢?(详见 Ridley 2000 中的深思熟虑的讨论)。
据说,生命本身取决于支持高保真信息流从一代传递到下一代的机制的进化。更有雄心壮志的是,梅纳德·史密斯和萨特玛里认为,在过去 40 亿年的进化中,许多关键步骤——他们所称的“进化中的重大转变”——涉及到创造新的跨代传递信息的方式——更可靠、更精细、更强大的方式,使得生物复制事件中的形态可靠地再现。从基于 RNA 的遗传系统转变为基于 DNA 的遗传系统(可能)是一个中心例子。但梅纳德·史密斯和萨特玛里认为,从大猿形式的社会生活转变为人类社会生活是一个重大转变,部分原因是因为人类社会生活具有新颖的大规模合作形式,但更主要的是因为他们将人类语言视为突破性的信息技术,彻底改变了高保真代际文化学习的可能性(MacArthur 1958; Maynard Smith and Szathmáry 1995, 1999)。
Maynard Smith 和 Szathmáry 关于重大转变的工作在宏观进化思维中具有里程碑意义-思考生命历史中的大规模模式。但是他们的工作中的信息维度并未被采纳。一个小的例外是 Sterelny(2009)。这篇论文认为,多细胞动物生命不仅依赖于高保真度的更多信息传递,还依赖于在发育中对该信息的控制,这表明卵的进化-一个受控的、结构化的、信息丰富的发育环境-对于复杂的动物生命至关重要。但是大多数关注点都集中在他们关于重大转变的另一个方面:解决合作问题,然后使先前独立的个体组合成新的、更复杂的个体。因此,在 Calcott 和 Sterelny(2011)中,没有一篇论文主要关注代际信息流的扩展或控制。这可能会改变。遗传机制的进化关键特征现在常常以信息术语进行讨论,而语言和 DNA 中的组合结构为类比推理提供了有力的基础。
Bibliography
Austin, C., 2015, “The Dispositional Genome: Primus inter pares”, Biology and Philosophy, 30: 227–246
Bergstrom, C. and M. Rosvall, 2011, “The Transmission sense of Information”, Biology and Philosophy, 26: 159–176.
Bromham, L., 2008, Reading the Story in DNA: A Beginner’s Guide to Molecular Evolution, Oxford: Oxford University Press.
Calcott, B., 2014, “The Creation and Reuse of Information in Gene Regulatory Networks”, Philosophy of Science, 81: 1–12.
Calcott, B. and K. Sterelny (eds.), 2011, The Major Transitions in Evolution Revisited, Cambridge, MA: MIT Press.
Crick, F., 1958, “On Protein Synthesis”, Symposia of the Society for Experimental Biology, 12: 138–163.
Dawkins, R., 1995, River out of Eden: A Darwinian View of Life, New York: Basic Books.
Francis, R., 2003, Why Men Won’t Ask for Directions: The Seductions of Sociobiology, Princeton: Princeton University Press.
Frank, S., 2012, “How to Read the Fundamental Equations of Evolutionary Change in Terms of Information Theory”, Journal of Evolutionary Biology, 25: 2377–2396.
Gilbert, S., 2013, Developmental Biology, Sunderland: Sinauer.
Godfrey-Smith, P., 1999, “Genes and Codes: Lessons from the Philosophy of Mind?”, in Biology Meets Psychology: Constraints, Conjectures, Connections, V. Hardcastle (ed.), Cambridge, MA: MIT Press.
–––, 2000, “On the Theoretical Role of Genetic Coding”, Philosophy of Science, 67: 26–44.
–––, 2007, “Information in Biology”, in The Cambridge Companion to the Philosophy of Biology, D. Hull and M. Ruse (eds.), Cambridge: Cambridge University Press.
–––, forthcoming, “Sender-Receiver Systems Within and Between Organisms”, Philosophy of Science.
Gray, R.D., 2001, “Selfish Genes or Developmental Systems?”, in Thinking about Evolution: Historical, Philosophical and Political Perspectives, R. Singh, K. Krimbas, D. Paul and J. Beatty (eds.), Cambridge: Cambridge University Press, 184–207.
Grice, H.P., 1957, “Meaning”, Philosophical Review, 66: 377–388.
Griesemer, J., 2005, “The Informational Gene and the Substantial Body: On the Generalization of Evolutionary Theory By Abstraction”, in Idealization and Abstraction in the Sciences, N. Cartwright and M. Jones (eds.), Amsterdam/New York: Editions Rodopi. XII.
Griffiths, P.E., 2001, “Genetic Information: A Metaphor in Search of a Theory”, Philosophy of Science, 68: 394–412.
Griffiths, P.E. and R.D. Gray, 1994, “Developmental Systems and Evolutionary Explanation”, Journal of Philosophy, XCI(6): 277–304.
–––, 2001, “Darwinism and Developmental Systems”, in Cycles of Contingency: Developmental Systems and Evolution, S. Oyama, P. Griffiths and R. Gray (eds.), Cambridge, MA: MIT Press, 195–218.
–––, 2004, “The Developmental Systems Perspective: Organism-environment Systems as Units of Evolution”, in The Evolutionary Biology of Complex Phenotypes, K. Preston and M. Pigliucci (eds.), Oxford and New York: Oxford University Press, 409–431.
Griffiths, P.E. and E. Neumann-Held, 1999, “The Many Faces of the Gene”, BioScience, 49(8): 656–662.
Griffiths, P.E. and K. Stotz, 2013, Genetics and Philosophy: An Introduction, Oxford: Oxford University Press.
Haig, D., 1997, “Parental Antagonism, Relatedness asymmetries, and Genomic Imprinting”, Proceedings of the Royal Society of London B, 264: 1657–1662.
Jablonka, E., 2002, “Information: its Interpretation, its Inheritance, and its Sharing”, Philosophy of Science, 69(4): 578–605.
Jablonka, E. and M. Lamb, 2005, Evolution in Four Dimensions, Cambridge, MA: MIT Press.
Jablonka, E. and E. Szathmáry, 1995, “The Evolution of Information Storage and Heredity”, Trends in Ecology and Evolution, 10(5): 206–211.
Kay, L., 2000, Who Wrote the Book of Life? A History of the Genetic Code, Palo Alto, CA: Stanford University Press.
Kirschner, M. and J. Gerhart, 1998, “Evolvability”, Proceedings of the National Academy of Science, USA, 95: 8420–8427.
Kirschner, M.W. and J.C. Gerhart, 2005, The Plausibility of Life: Resolving Darwin’s Dilemma, New Haven: Yale University Press.
Knight, R., S. Freeland, and L. Landweber, 1999, “Selection, History and Chemistry: The Three Faces of the Genetic Code”, Trends in Biochemical Sciences, 24: 241–247.
Krebs, J. and R. Dawkins, 1984, “Animal Signals, Mind-Reading and Manipulation”, in Behavioural Ecology: An Evolutionary Approach, J.R. Krebs and N.B. Davies (eds.), Oxford: Blackwell Scientific: 380–402.
Lean, O., 2014, “Getting the Most out of Shannon Information”, Biology and Philosophy, 29: 395–413.
Lehrman, D.S., 1970, “Semantic and Conceptual Issues in the Nature-Nurture Problem”, in Development and Evolution of Behaviour, D.S. Lehrman (ed.), San Francisco: W.H. Freeman.
Levy, A., 2011, “Information in Biology: A Fictionalist Account”, Noûs, 45(4): 640–657.
Lewontin, R.C., 1974, “The Analysis of Variance and the Analysis of Causes”, American Journal of Human Genetics, 26: 400–411.
–––, 2000, The Triple Helix, Cambridge, MA: Harvard University Press.
Lorenz, K., 1965, Evolution and The Modification of Behaviour, Chicago: Chicago University Press.
MacArthur, R.H., 1958, “Population Ecology of Some Warblers of Northeastern Coniferous Forests”, Ecology, 30: 599–619.
Maclaurin, J., 1998, “Reinventing Molecular Weissmanism”, Biology and Philosophy, 13(1): 37–59.
Maynard Smith, J., 2000, “The Concept of Information in Biology”, Philosophy of Science, 67(2): 177–194.
Maynard Smith, J. and E. Szathmáry, 1995, The Major Transitions in Evolution, New York: Freeman.
–––, 1999, The Origins of Life: From the Birth of Life to the Origins of Language, Oxford: Oxford University Press.
Millikan, R., 1984, Language, Thought and Other Biological Categories, Cambridge, MA: MIT Press.
Owren, M.J., D. Rendall, and M.J. Ryan, 2010, “Redefining Animal Signaling: Influence versus Information in Communication”, Biology and Philosophy, 25(5): 755–780.
Oyama, S., 1985/2000, The Ontogeny of Information, Durham: Duke University Press.
Planer, R., 2014, “Replacement of the ‘Genetic Program’ Program”, Biology and Philosophy, 29(1): 33–53.
Ridley, M., 2000, Mendel’s Demon: Gene Justice and the Complexity of Life, London: Weidenfeld and Nicholson.
Sansom, R., 2008a, “The Connectionist Framework for Gene Regulation”, Biology & Philosophy, 23(4): 475–491.
–––, 2008b, “Countering Kauffman with Connectionism: Two Views of Gene Regulation and the Fundamental Nature of Ontogeny”, The British Journal for the Philosophy of Science, 59(2): 169–200.
–––, 2011, Ingenious Genes: How Gene Regulation Networks Evolve to Control Development, Cambridge, MA: MIT Press.
Sarkar, S., 1996, “Decoding ‘Coding’—Information and DNA”, Biosciences, 46: 857–864.
–––, 2003, “Genes Encode Information for Phenotypic Traits”, in Contemporary Debates in Philosophy of Science, C. Hitchcock (ed.), London, Blackwell: 259–272.
Schrödinger, E., 1944, What is Life?, Cambridge: Cambridge University Press.
Shannon, C., 1948, “A Mathematical Theory of Communication”, Bell Systems Technical Journal, 27: 279–423, 623–656.
Shea, N., 2007a, “Consumers Need Information: Supplementing Teleosemantics with an Input Condition”, Philosophy and Phenomenological Research, 75: 404–435.
–––, 2007b, “Representation in the genome and in other inheritance systems”, Biology and Philosophy, 22(3): 313–331.
–––, 2011, “Developmental Systems Theory Formulated as a Claim about Inherited Representations”, Philosophy of Science, 78(1): 60–82.
–––, 2013, “Inherited Representations are Read in Development”, British Journal for the Philosophy of Science, 64(1): 1–31.
Skyrms, B., 2010, Signals: Evolution, Learning and Information, Oxford: Oxford University Press.
Stegmann, U., 2004, “The Arbitrariness of the Genetic Code”, Biology and Philosophy, 19: 205–222.
–––, 2012, “Varieties of Parity”, Biology and Philosophy, 27: 903–918.
–––, 2014, “Causal Control and Genetic Causation”, Noûs, 48(3): 450–465.
Sterelny, K., 2004, “Symbiosis, Evolvability and Modularity”, in Modularity in Development and Evolution, G. Schlosser and G. Wagner (eds.), Chicago: University of Chicago Press, 490–516.
–––, 2009, “Novelty, Plasticity and Niche Construction: The Influence of Phenotypic Variation on Evolution”, in Mapping the Future of Biology: Evolving Concepts and Theories, A. Barberousse, M. Morange and T. Pradeu (eds.), Dordrecht: Springer.
–––, 2011, “Evolvablity Revisited”, in The Major Transitions Revisited, B. Calcott and K. Sterelny (eds.), Cambridge, MA: MIT Press, 83–100.
Sterelny, K., K.C. Smith, and M. Dickison, 1996, “The Extended Replicator”, Biology and Philosophy, 11: 377–403.
Szathmáry, E. and J. Maynard Smith, 1997, “From Replicators to Reproducers: The First Major Transitions Leading to Life”, Journal of Theoretical Biology, 187: 555–571.
Waters, C., K, 2007, “Causes That Make a Difference”, Journal of Philosophy, 104: 551–579.
Williams, G.C., 1992, Natural Selection: Domains, Levels and Challenges, Oxford: Oxford University Press.
Woodward, J., 2010, “Causation in Biology: Stability, Specificity, and the Choice of Levels of Explanation”, Biology and Philosophy, 25: 25:287–318.
Wright, L., 1973, “Functions”, The Philosophical Review, 82: 139–168.
Academic Tools
Look up topics and thinkers related to this entry at the Internet Philosophy Ontology Project (InPhO). | |
Enhanced bibliography for this entry at PhilPapers, with links to its database. |
Other Internet Resources
[Please contact the author with suggestions.]
Related Entries
functionalism | genetics: ecological | genetics: genotype/phenotype distinction | heritability | mental content: teleological theories of | mental representation | naturalism | natural selection | teleology: teleological notions in biology
最后更新于

{kind=link}